探讨瘦素受体(LEPR)基因表达及其多态性与乳腺癌发生、发展及病理学特征之间的关系。
采用免疫组织化学法和聚合酶链反应-限制性片段长度多态(PCR-RFLP)分析技术,对150例乳腺癌患者肿瘤组织、80例乳腺良性病变患者病变组织、50例患者癌旁正常乳腺组织和128例健康对照者静脉血进行LEPR基因表达及Gln223Arg多态性检测。
LEPR在乳腺癌组织中表达的阳性率为70.67 %(106/150),高于乳腺良性病变组的56.25 %(45/80)和癌旁正常组织组的44.00 % (22/50)(均P<0.005)。LEPR基因Gln223Arg的GG、GA、AA基因型在乳腺癌组的分布频率分别为70.00 %(105例)、16.67 %(25例)和13.33 %(20例),在良性病变组为82.5 %(66例)、13.75 %(11例)、3.75 %(3例),在癌旁正常组织组为82.00 %(41例)、14.00 %(7例)、4.00 %(2例),在健康对照组为82.81 %(106例)、14.85 %(19例)和2.34 %(3例),三组比较差异有统计学意义(χ2=11.56,P= 0.003),GG、GA、AA基因型在乳腺良性疾病组、癌旁正常组织组和健康对照组分布频率比较,差异无统计学意义(P>0.05)。乳腺癌组G和A等位基因分布频率分别为78.33 %(235例)和21.67 %(65例),与良性病变组、癌旁正常组织组及健康对照组相比,差异有统计学意义(χ2=12.52, P=0.001)。LEPR基因在淋巴结转移患者中阳性表达率为87.8 %,高于无淋巴结转移患者的60.2 %,差异有统计学意义(P= 0.02)。多因素分析表明,LEPR基因高表达、Gln223Arg多态性及腰臀比(WHR)升高是乳腺癌发生的危险因素(均OR>1)。
LEPR高表达、Gln223Arg多态性及WHR升高可能增加乳腺癌发生的风险。


版权归中华医学会所有。
未经授权,不得转载、摘编本刊文章,不得使用本刊的版式设计。
除非特别声明,本刊刊出的所有文章不代表中华医学会和本刊编委会的观点。
近年来,随着脂肪组织内分泌功能及其分泌的多种肽类物质的发现,人们对肥胖促进乳腺癌发生的机制有了新的认识,认为肥胖是乳腺癌发生的一个独立危险因素[1,2,3]。瘦素(LEP)作为一种完全由脂肪组织生成的神经内分泌激素,可与其受体结合发挥生物学作用,可能参与了乳腺癌的发生。我们对乳腺癌、乳腺良性病变和癌旁正常乳腺组织中瘦素受体(LEPR)基因的表达及其单核苷酸多态性(SNP)进行检测,以探讨LEPR基因在乳腺癌发生、发展中的作用。
选择山西省肿瘤医院乳腺治疗中心2009年1月至2010年10月收治的原发性乳腺癌患者150例(其中浸润性导管癌68例,导管原位癌59例,浸润性小叶癌11例,小叶原位癌12例),乳腺良性病变80例(纤维腺瘤47例,多发性导管内乳头状瘤17例,小叶增生16例);健康对照128例。收集乳腺癌、乳腺良性病变组织及50例距癌组织4 cm处的正常乳腺组织标本,离体时间<4 h,均用4%中性甲醛溶液固定24 h,常规石蜡包埋,采用免疫组织化学法测定LEPR表达。同时取乳腺癌、乳腺良性病变新鲜组织0.5~ 1.0 g,经0.9 % NaCl溶液冲洗后采用酚、氯仿法提取DNA,健康对照抽取肘静脉抗凝血,采用高盐沉淀法提取外周血白细胞基因组DNA,-28 ℃保存准备行LEPR基因Gln223Arg多态性检测。对所有入组者年龄、绝经状态等基本资料进行登记,并测量身高、体质量、腰围和臀围,计算体质量指数(BMI)和腰臀比(WHR)。
LEPR兔抗人多克隆抗体(M-18)(美国Santa Cruz公司);快速免疫组织化学EnVision检测试剂盒、KIT-5004和二氨基联苯胺(DAB)显色试剂盒均由福州迈新生物技术开发公司提供。PCR扩增试剂盒、内切酶、引物序列合成等均由上海生工生物工程有限公司提供。9700型荧光定量PCR仪为美国PE公司产品,22331 Hamburg DNA浓度测定仪为德国Eppendorf公司产品,Smart View 2001生物电泳图像分析仪由上海复日公司提供,Sunrise酶标仪为奥地利Tecan公司产品。
采用组织芯片制备机和纯净石蜡等将各组乳腺组织标本制成组织芯片,将普通蜡块与制好的组织芯片4 μm厚连续切片,60 ℃烤片4 h。采用即用型EnVision两步法,脱蜡,水化,高压热修复,H2O2封闭内源性过氧化物酶活性。分别滴加LEPR一抗,4 ℃冰箱过夜,滴加EnVision,DAB显色,苏木精对比染色。脱水,透明,封固,镜检。
LEPR为胞质和胞膜表达,内有黄色或棕黄色为阳性。光学显微镜下观察5个高倍视野,计算染色阳性细胞所占的比例。-:阴性细胞数<10%;+:阳性细胞数为10%~50%;++:阳性细胞数>50 %,染色弱;+++:阳性细胞数>50 %,染色强。其中++和+++都为强阳性染色。
2×HiFi-PCR Master 25 μl加上游引物(5'-ACCC TTTAAGCTGGGTGTCCCAAATGA-3')2 μl,加下游引物(5'-GCTAGCAAATATTTTTGTAAGCAATT-3')2 μl,加DNA模板10 μl,加Sterial ddH2O 11 μl共50 μl。反应体系:94 ℃预变性5 min后进行30个循环,94 ℃变性60 s, 55 ℃复性60 s, 72 ℃延伸60 s,循环结束后72 ℃终延伸5 min。取10 μl PCR扩增产物与2 μl 6×上样缓冲液混匀,在2.0%琼脂糖凝胶中上样、电泳,电压为100 V,时间为30~ 40 min,以Marker为参照,在紫外灯下观察扩增产物片断为440 bp。
酶切反应体系,取PCR扩增产物10 μl加10×酶切缓冲液15 μl加2 μl内切酶(MspⅠ)加无菌去离子水23 μl共50 μl,在37 ℃中放置过夜。取10 μl PCR酶切产物加2 μl 6×上样缓冲液混匀,在2.0 %琼脂糖凝胶中上样、电泳,电压为100 V,时间为30~ 40 min,以Marker为参照,在紫外灯下观察三种不同基因型结果,Gln223纯合子(AA)电泳图谱为440 bp一条带;Arg223纯合子(GG)电泳图谱为300 bp和140 bp两条带,Gln223/Arg223杂合子(GA)电泳图谱为300 bp、140 bp和440 bp三条带。
使用SPSS 13.0统计软件进行数据分析,计量资料两组间采用t检验比较;计数资料比较采用χ2检验,其他因素的比较采用相关和非条件Logistic回归分析等。以P<0.05为差异有统计学意义。
乳腺癌组、乳腺良性病变组及健康对照组在年龄、职业、种族、地域、文化程度方面匹配良好。BMI在乳腺癌组为25.05 kg/m2,高于乳腺良性病变组的23.36 kg/m2和健康对照组的23.07 kg/m2,差异有统计学意义(P<0.005);乳腺癌组WHR为0.86,高于乳腺良性病变组的0.82,差异无统计学意义(P= 0.24)。
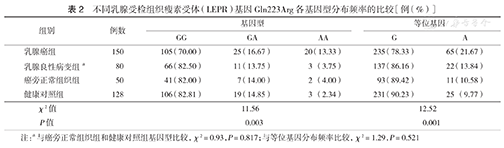
LEPR基因在乳腺癌组、良性病变组和癌旁正常组织组表达的阳性率分别为70.67 %、56.25%和44.00 %;强阳性率分别为32.00 %、22.50%和4.00 %。乳腺癌组阳性率较其他两组高,差异均有统计学意义(均P<0.05 ),乳腺良性病变组与癌旁正常组织组比较差异无统计学意义(P>0.05)(表1,图1, 图2, 图3)。
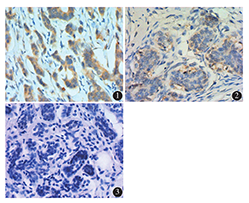
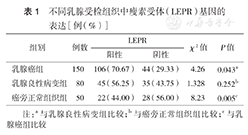
不同乳腺受检组织中瘦素受体(LEPR)基因的表达[例(%)]
不同乳腺受检组织中瘦素受体(LEPR)基因的表达[例(%)]
| 组别 | 例数 | LEPR | χ2值 | P值 | |
|---|---|---|---|---|---|
| 阳性 | 阴性 | ||||
| 乳腺癌组 | 150 | 106(70.67) | 44(29.33) | 4.26 | 0.043a |
| 乳腺良性病变组 | 80 | 45(56.25) | 35(43.75) | 1.328 | 0.252b |
| 癌旁正常组织组 | 50 | 22(44.00) | 28(56.00) | 8.23 | 0.005c |
注:a与乳腺良性病变组比较;b与癌旁正常组织组比较;c与乳腺癌组比较
平衡性检验结果表明,健康对照组LEPR基因Gln223Arg的GG、GA、AA基因型分布频率分别为82.81 %、14.85%和2.34 %,与理论分布的78.83 %、19.82%和1.35%比较,差异无统计学意义(P>0.05);等位基因G和A分布频率为90.23%和9.77 %,与理论分布的88.9%和11.1%比较,差异无统计学意义(P>0.05 )。基因型分布频率符合Hardy-Weinberg平衡定律,提示所研究的样本具有群体代表性。
LEPR基因Gln223Arg的GG、GA、AA基因型在乳腺癌组、乳腺良性病变组、癌旁正常组织组、健康对照组的分布频率比较,差异有统计学意义(χ2= 11.56, P=0.003 )。三组间G和A等位基因分布频率比较,差异亦有统计学意义(χ2=12.52,P=0.001)。而良性病变组、癌旁正常组织组和健康对照组基因型与各等位基因的分布频率差异均无统计学意义(均P>0.05)(表2)。
不同乳腺受检组织瘦素受体(LEPR)基因Gln223Arg各基因型分布频率的比较[例(%)]
不同乳腺受检组织瘦素受体(LEPR)基因Gln223Arg各基因型分布频率的比较[例(%)]
| 组别 | 例数 | 基因型 | 等位基因 | ||||
|---|---|---|---|---|---|---|---|
| GG | GA | AA | G | A | |||
| 乳腺癌组 | 150 | 105(70.00) | 25(16.67) | 20(13.33) | 235(78.33) | 65(21.67) | |
| 乳腺良性病变组a | 80 | 66(82.50) | 11(13.75) | 3(3.75) | 137(86.16) | 22(13.84) | |
| 癌旁正常组织组 | 50 | 41(82.00) | 7(14.00) | 2(4.00) | 93(89.42) | 11(10.58) | |
| 健康对照组 | 128 | 106(82.81) | 19(14.85) | 3(2.34) | 231(90.23) | 25(9.77) | |
| χ2值 | 11.56 | 12.52 | |||||
| P值 | 0.003 | 0.001 | |||||
注:a与癌旁正常组织组和健康对照组基因型比较,χ2=0.93, P=0.817;与等位基因分布频率比较,χ2=1.29, P=0.521
携带AA基因型和GA基因型与携带GG基因型者相比,患乳腺癌的OR分别为7.25和1.28,携带AA+GA型者患乳腺癌的OR为2.13(表3)。
不同乳腺受检组织瘦素受体基因Gln223Arg多态性与乳腺癌发生的关系[例(%)]
不同乳腺受检组织瘦素受体基因Gln223Arg多态性与乳腺癌发生的关系[例(%)]
| 组别 | 例数 | 基因型别 | ||||||
|---|---|---|---|---|---|---|---|---|
| GG | GA | AA | GA+AA | |||||
| 乳腺癌组 | 150 | 105(70.67) | 25(16.67) | 20(13.33) | 45(30.00) | |||
| 乳腺良性病变组 | 80 | 66(82.50) | 11(13.75) | 3(3.75) | 14(17.50) | |||
| 健康对照组 | 128 | 106(82.81) | 19(14.85) | 3(2.34) | 22(17.19) | |||
| 危险度(OR) | 1.00 | 1.28 | 7.25 | 2.13 | ||||
| 95 % CI | 1.05~2.57 | 1.90~26.2 | 1.07~3.22 | |||||
| P值 | 0.030 | 0.001 | 0.005 | |||||
LEPR基因的表达与乳腺癌患者绝经状态、组织学类型、肿瘤大小、肿瘤分级和远处转移状况等均无相关性(均P>0.05),与雌激素受体(ER)和孕激素受体(PR)的表达状况也无相关性(P>0.05 )。但LEPR基因在淋巴结转移患者中的阳性表达率为87.8 %,高于无淋巴结转移患者的60.2 %,二者差异有统计学意义(P=0.02)。
将单因素分析有意义的因素和可能的危险因素引入非条件Logistic回归模型,采用逐步回归法进行多因素分析(α=0.05,β=0.10),最终进入方程的有LEPR基因表达、Gln223Arg多态性和WHR。结果显示,LEPR基因表达、Gln223Arg多态性及WHR的OR均大于1,它们可能是乳腺癌发生的危险因素(表4)。
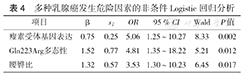
多种乳腺癌发生危险因素的非条件Logistic回归分析
多种乳腺癌发生危险因素的非条件Logistic回归分析
| 项目 | β | s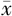 | OR | 95 % CI | Wald | P值 |
|---|---|---|---|---|---|---|
| 瘦素受体基因表达 | 0.75 | 0.25 | 5.06 | 1.25~10.27 | 8.33 | 0.002 |
| Gln223Arg多态性 | 1.52 | 0.77 | 4.81 | 1.35~18.22 | 5.21 | 0.012 |
| 腰臀比 | 1.32 | 0.57 | 3.53 | 1.30~10.23 | 6.45 | 0.017 |
LEP通过与LEPR结合发挥其生物学作用,是经典Ⅰ类细胞因子受体家族的成员之一。LEP作用于LEPR,对机体代谢起着调节作用。LEPR胞内端含有三个关键的酪氨酸位点,其磷酸化能够激活JAK2-STAT3及ERK等通路,以此控制下游的信号通路,进而调节能量摄取与能量消耗的平衡以及葡萄糖的代谢。上述代谢过程紊乱是引起肥胖及肥胖相关疾病的直接原因。Tartaglia等[4]证实小鼠LEPR基因定位于4号染色体,而人类LEPR基因位于1p31,长约5.1 kb,含20个外显子,19个内含子,编码1 165个氨基酸。LEPR是LEP的高亲和力受体,属Ⅰ类细胞因子超家族。LEPR有6种亚型,即可溶性受体(OB-Re)、长型受体(OB-Rb)和4种短型受体(OB-Ra、OB-Rc、OB-Rd、OB-Rf),其中OB-Rb被认为是其功能受体。LEP和LEPR广泛存在于人体各种组织中,如下丘脑、脂肪组织、神经、心脏、肾脏、乳腺、肺、肝脏及胰岛细胞表面等,LEP与其受体结合产生生物学作用。有研究发现LEPR基因序列中的单碱基变异体多态性可导致基因产物变化,并与多种代谢性疾病相关,如肥胖、糖尿病、肿瘤及代谢综合征等[5,6,7]。
Ishikawa等[8]采用特异性抗体免疫组织化学方法,检查76例乳腺浸润性导管癌和32例癌旁正常乳腺组织中LEP和LEPR基因的表达情况,发现正常乳腺上皮并不显著表达LEPR,而有63例(83 %)在癌细胞中LEPR呈现阳性染色。癌细胞和乳腺正常上皮细胞均表达LEP,在乳腺癌患者中有70例(92 %) LEP过表达,而在正常上皮中则没有过表达现象。Garofalo等[9]对148例乳腺癌组织和90例乳腺良性病变组织进行研究发现,乳腺癌组织LEPR表达率为41.2 %,而乳腺良性病变中未发现LEPR的表达。Revillion等[10]分析了322例原发性乳腺癌中LEP、LEPR-L和LEPR-S的表达情况,发现所有的肿瘤均表达两种受体,其中318例表达LEP。三个变量之间有关联性,并与雌二醇及孕酮受体相关,但与组织恶性程度和肿瘤大小无关。本研究结果发现LEPR基因在乳腺癌组织中广泛表达,阳性率为70.67 %,提示LEPR在乳腺癌的发生和发展中起重要的作用,LEPR在乳腺癌组织中的高表达会使乳腺癌细胞更易接受LEP的应答。
Garofalo等[9]发现在有淋巴结转移的乳腺癌组织中LEPR的表达率为51.5%,明显高于未发生淋巴结转移乳腺癌组织的41.2 %。本研究发现LEPR基因在淋巴结转移组阳性率(87.8 %)明显高于无淋巴结转移患者(60.2 %),提示LEPR参与了乳腺癌组织的增殖过程,对于乳腺癌的发展起着非常重要的作用。本研究结果也发现LEPR基因的表达与患者年龄、绝经状态、肿瘤大小、肿瘤病理学特征、ER、PR均无关,与Ishikawa等[8]研究结果一致。
LEPR基因的多态性来自于基因序列中的单碱基变异,迄今为止,已经发现多种LEPR基因多态性,这些多态性导致其基因产物产生变化,它们的变化与多种疾病相关,如肥胖、糖尿病等。1997年Matsuoka等[11]对LEPR基因的2 ~20外显子应用PCR技术进行了测序分析,发现了7种核苷酸变异。我们的研究结果也表明,LEPR基因Gln223Arg位点多态性的分布频率在乳腺癌与乳腺良性病变、癌旁正常组织和健康对照组差异有统计学意义,提示LEPR基因Gln223Arg位点多态性可能与乳腺癌的发生有关。到目前为止,关于LEPR基因Gln223Arg多态性与乳腺癌关系的研究甚少。Woo等[12]的研究认为LEPR基因Gln223Arg多态性与乳腺癌的发生无关。Ishikawa等[8]研究结果表明,LEPR基因变异可能是乳腺癌发生的危险因素。关于LEPR基因Gln223Arg多态性在乳腺癌发生、发展过程中产生的作用有待于进一步研究。
近年来,流行病学调查结果表明,超重、肥胖、营养过剩及代谢紊乱等与乳腺癌的发生有关。O'Brien等[13]报道,在欧美一些国家,随着中老年妇女体质量增加,乳腺癌的发病率上升。Borugian等[14]研究结果表明,升高的BMI、WHR为妇女患乳腺癌的重要危险因素。本研究结果进一步证实,乳腺癌患者BMI、WHR明显高于乳腺良性病变和健康对照者,多因素分析表明,LEPR基因表达、Gln223Arg多态性及WHR OR值均大于1,它们可能是乳腺癌发生的危险因素。Kuriyama等[15]报道肥胖妇女的LEP水平上升有可能促进乳腺癌的发生,BMI是增加绝经后妇女乳腺癌发生的主要危险因素,与绝经前妇女无关,同时也发现绝经前后妇女患乳腺癌的风险与体力活动有明显的相关性。

























