
探究在脂多糖(lipopolysaccharide ,LPS )诱导的大鼠膀胱炎中核因子-κB (nuclear factor kappa B ,NF-κB)信号通路的激活对膀胱中胶原蛋白分泌的影响。
选用体重200~230 g的健康雌性SD大鼠48只,按数字随机法随机分为空白对照组(A组)、假手术组(B组)、LPS灌注2周组(C组)、LPS灌注4周组(D组)、抑制剂对照组(E组)和抑制剂组(F组),每组8只。用LPS对C、D、E、F组大鼠进行反复灌注诱导大鼠膀胱炎的发生,用二硫代氨基甲酸吡咯烷(pyrrolidin-edithiocarbamic acid ,PDTC)作为NF-κB抑制剂。所有大鼠处死前均进行6 h内的排尿次数记录。对膀胱标本进行HE染色评估膀胱的炎症情况,再通过免疫组化染色检测膀胱组织中NF-κB p65、磷酸化p65、Col1和Col3A1蛋白的含量表达。
LPS灌注组大鼠膀胱的HE染色结果显示黏膜下层水肿、黏膜上皮增生、糜烂,炎症细胞浸润明显。LPS灌注组中膀胱上皮中Col1和Col3A1蛋白的表达均较对照组升高,差异有统计学意义(P<0.05)。NF-κB p65及磷酸化p65在LPS灌注组中的表达也均较空白对照组及pH7.4磷酸缓冲盐溶液(phosphate buffer saline,PBS)灌注组升高,差异有统计学意义(P<0.05 )。在抑制剂组中,大鼠膀胱中NF-κB p65的活化受到了抑制;同时,大鼠膀胱的炎症浸润及尿频症状均比对照组有所改善;而在胶原蛋白表达方面,抑制剂组膀胱上皮中Col1蛋白的含量比对照组降低,差异有统计学意义(P<0.05),Col3A1蛋白的含量未见降低,差异无统计学意义(P> 0.05)。
NF-κB信号通路为大鼠膀胱炎症刺激膀胱胶原沉积过程中的重要信号通路,该通路的激活可能是纤维化过程的重要因素;而PDTC作为一种NF-κB抑制剂,不仅能有效改善大鼠的膀胱炎症情况及尿频症状,还能一定程度上抑制膀胱上皮细胞的胶原蛋白的合成分泌,延缓纤维化进程。


版权归中华医学会所有。
未经授权,不得转载、摘编本刊文章,不得使用本刊的版式设计。
除非特别声明,本刊刊出的所有文章不代表中华医学会和本刊编委会的观点。
革兰阴性菌作为临床上泌尿系感染的主要致病菌,其主要致病因子为脂多糖(lipopolysaccharide , LPS)。LPS作为革兰阴性菌主要的致病因子,可通过Toll样受体4(TLR4)导致NF-κB通路的活化,产生大量的促炎症因子,诱导炎症进程的发生发展[1,2]。革兰阴性菌在膀胱中的种植感染易致患儿反复的膀胱炎症,尤其是临床上存在下尿路功能障碍的患儿,如神经源性膀胱患儿,容易出现反复的泌尿系感染,这种反复的泌尿系感染不仅治疗棘手,且严重影响患儿的生活质量,长期以往最终易致膀胱纤维化的发生。膀胱纤维化是以细胞外基质(extracellular matrix, ECM)在细胞间质、肌层间异常增多并过度沉积而导致膀胱组织结构破坏和功能减退为特征的病理过程。目前由于纤维化的具体发生机制目前仍不清楚,因此治疗也仍是难题。
核因子-κB (nuclear factor kappa B ,NF-κB)是真核细胞内一类重要的转录因子,几乎在所有类型的细胞中该转录因子均有表达;与炎症反应、免疫反应以及细胞增殖、分化、凋亡等密切相关[3]。NF-κB通路作为诱导炎症反应发生和发展的重要通路,既往研究已证实其能诱导膀胱的炎性改变[4,5],也证实了其在肾脏、肝脏等其他器官中促进纤维化的发生进程中的重要作用[6,7,8]。二硫代氨基甲酸吡咯烷(pyrrolidinedithiocarbamic acid ,PDTC)作为一种抗氧化剂,能抑制NF-κB抑制蛋白IκB (inhibitor kappa B ,IκB )的降解,进而抑制NF-κB的激活。Okabe等[9]证实,PDTC能通过减轻肾脏炎症反应进而减轻肾脏纤维化的发生。该通路与膀胱纤维化之间的关系如何,目前相关研究较少,这也是本研究的主要目的。
本研究拟通过效仿前人诱导大鼠膀胱炎的建模方法[4,5],使用LPS对大鼠进行膀胱灌注诱导膀胱炎症,通过LPS反复刺激,观察大鼠膀胱炎症及纤维化的情况。在此过程中,同时检测NF-κB通路相关信号分子在膀胱组织中的表达情况,以探究NF-κB通路在LPS刺激所致的膀胱炎症到膀胱发生纤维化进程中的表达变化。然后,再使用PDTC作为NF-κB通路抑制剂,通过阻断NF-κB通路的激活,再观察膀胱中胶原蛋白的沉积情况,以进一步观察NF-κB通路与膀胱纤维化之间的关系。为研究膀胱纤维化的发生机制及治疗提供合适的靶点参考。
LPS(美国CST公司);NF-κB p65兔多抗(美国CST公司);磷酸化NF-κB p65兔多抗(德国Gentex);Col1兔多抗(Novus公司);Col3A1兔多抗(Novus );免疫组化试剂盒(英国Abcam公司);硬膜外麻醉导管(驼人医疗)。深圳市儿童医院本院病理科进行HE染色。
健康的SD大鼠,SPF级,雌鼠48只,体重200~230 g,实验动物来自南方医科大学实验动物中心[许可证号:SYXK(粤)2018-0186]。按数字随机法随机分为空白对照组(A组)、pH7.4磷酸缓冲盐溶液(phosphate buffer saline ,PBS)灌注组(B组)、LPS灌注2周(C组)、4周(D组)、抑制剂对照组(E组)和抑制剂组(F组),每组8只。本实验符合国家实验动物福利伦理的相关规定(伦理编号IACUC-G16026)并遵守赫尔辛基宣言。
采用10%水合氯醛溶液0.3 ml/100g腹腔注射对大鼠进行麻醉,麻醉生效后,将大鼠仰卧位固定于操作台上。将大鼠会阴部进行剃毛备皮,碘伏消毒会阴部。将硬膜外麻醉导管剪取前端一段长约10 cm以代替导尿管,消毒后自尿道外口导入膀胱,轻压腹部可见清亮尿液自导管内流出即证明导尿成功。导尿成功后,缓慢持续按压大鼠腹部让膀胱排空,然后将药物缓慢灌注导入膀胱。膀胱灌注完成后继续留置导尿管防止漏尿,40 min后缓慢拔除导尿管。隔天进行一次膀胱灌注,直至实验终点。
处死前1 d将大鼠置于代谢笼中,正常饲养,让其适应代谢笼环境。处死当天将大鼠取出,反复轻柔按压大鼠膀胱,直至无明显尿液排出。将大鼠放回代谢笼中,让其自由喝水。用摄像头记录此后6 h内大鼠的排尿次数。记录完成后再处死大鼠取膀胱标本。
A组:空白对照组,常规饲养4周。B组:假手术组,使用PBS缓冲液对大鼠进行膀胱灌注,隔天1次,持续4周。C组:使用LPS(使用PBS缓冲液稀释至1 mg/mL)对大鼠进行膀胱灌注,隔天1次,持续2周。D组:使用LPS对大鼠进行膀胱灌注,隔天1次,持续4周。E组:抑制剂对照组,使用LPS对大鼠进行膀胱灌注,隔天1次,持续4周;第5周停止灌注,改为使用注射灭菌用水进行腹腔注射,每天1次,连续1周。F组:抑制剂组,使用LPS对大鼠进行膀胱灌注,隔天1次,持续4周;第5周停止灌注,改为使用PDTC(灭菌注射用水稀释为50 mg/mL,剂量100 mg/kg)进行腹腔注射,每天1次,连续1周。所有大鼠在实验终点时记录完6h的排尿次数后再处死大鼠取膀胱标本。
取出膀胱标本后,用PBS浸洗将膀胱血迹清除,将标本浸泡于4%多聚甲醛固定48 h。而后进行脱水、石蜡包埋、切片。切片后分别进行HE染色及进行免疫组化染色。免疫组化染色按照试剂盒产品说明书进行操作:切片常规脱蜡复水后,pH6.0的柠檬酸钠抗原修复液使用微波炉加热法进行抗原修复;3%过氧化氢阻断,0.1%曲拉通进行穿透;10%山羊血清封闭1h;然后使用NF-κB p65 (1 :800)、NF-κB pp65 (1:50)、Col1 (1:150)和Col3A1(1:150)一抗孵育在4℃冰箱过夜;再进行二抗及链霉亲和素孵育;而后使用DAB显色,显微镜下观察组织显色情况,待特异性染色较强且背景着色较浅时即流水洗去DAB显色液;最后使用苏木精染色、脱水透明后进行封片。染色结果采用Image J软件对切片染色结果进行平均吸光度值比较分析。
对所收集的数据资料采用IBM SPSS 26.0统计学软件进行处理分析。计量资料用 ±s表示,多组比较及组间比较时采用单因素方差分析,两组间比较采用独立样本的t检验。P<0.05表示差异有统计学意义。
±s表示,多组比较及组间比较时采用单因素方差分析,两组间比较采用独立样本的t检验。P<0.05表示差异有统计学意义。
A、B、C组大鼠全部存活至实验终点;D组大鼠中途死亡1只,因麻醉不当,损伤肠管而死,其余大鼠均存活至实验终点。E组大鼠死亡1只,考虑腹腔感染而死,其余大鼠均存活至实验终点。F组大鼠全部存活至实验终点。
对HE染色病理切片进行镜下观察,结果显示,空白对照组及PBS灌注组膀胱上皮排列整齐、完整,未见明显炎症细胞浸润;而在LPS膀胱灌注2周后,黏膜下层水肿明显,膀胱上皮细胞部分排列紊乱,部分出现非典型性增生,可见较多中性粒细胞浸润黏膜下层;而在LPS灌注4周后,可见膀胱上皮细胞排列紊乱严重,黏膜上皮出现黏膜增生、糜烂,黏膜下层可见较多淋巴细胞浸润(图1)。

对每组大鼠实验终点时的6 h内的排尿次数进行了记录,结果如下:A、B、C、D组6 h的排尿频率分别为(2.88±0.78)次、(4.75±1.20)次、(9.50±2.69)次、(7.57±1.59)次。LPS膀胱灌注对大鼠的排尿频率的差异有统计学意义(F= 20.073,P<0.05);A组与B组比较,差异无统计学意义(P=0.129>0.05);C组、D组与B组进行组间比较,差异均有统计学意义(P<0.05)。
根据炎症的病理学诊断标准[10]:①变质;②渗出、水肿;③增生。结合LPS灌注组大鼠膀胱HE染色结果中黏膜下层的水肿、黏膜上皮的增生、糜烂,炎症细胞浸润以及下尿路刺激症状,证实大鼠膀胱炎模型已成功建立[4,5]。结果显示,在LPS膀胱灌注的2周左右,为大鼠膀胱炎的急性炎症期,此时可有明显的中性粒细胞浸润及炎症水肿;而在LPS膀胱灌注4周后,膀胱黏膜上皮开始出现糜烂,浸润的炎症细胞主要为淋巴细胞,提示着膀胱炎症有逐步进入慢性发展期的趋势。在下尿路刺激症状方面,B~F组大鼠对比空白对照组大鼠均出现了不同程度的尿频症状,且以LPS膀胱灌注的2周左右更为显著。
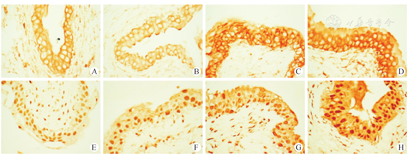
对组织切片进行Ⅰ型胶原蛋白(collagen Ⅰ,Col1)和Ⅲ型胶原蛋白(collagen Ⅲ ,Col3)的相关蛋白Col3A1的免疫组化染色,结果(分别见图2A-D,图2E-H)显示:两种胶原蛋白在对照组及LPS灌注组膀胱中均有表达,但在LPS膀胱灌注2周后表达明显升高,尤其是在膀胱上皮细胞中表达增加更加明显,而在LPS膀胱灌注4周后,两种胶原蛋白的分泌略有下降。本研究对膀胱上皮层的胶原蛋白表达情况用Image J软件对切片染色结果进行AOD值检测后再进行统计学分析。结果提示,对于Col1蛋白,LPS膀胱灌注对大鼠膀胱上皮Col1蛋白分泌增加的差异有统计学意义(F=200.198,P<0.05);A组与B组比较(0.233±0.015和0.243±0.013),差异无统计学意义(P=0.476>0.05);C组与B组比较(0.386±0.019和0.243±0.013)、D组与B组比较(0.368±0.011和0.243±0.013),差异均有统计学意义(P<0.05)。对于Col3A1蛋白,LPS膀胱灌注对大鼠膀胱上皮Col3A1蛋白分泌增加的差异也有统计学意义(F=135.319,P<0.05 ),空白对照组与PBS灌注组比较(0.213±0.012和0.220±0.012),差异无统计学意义(P=0.777>0.05),C组与B组比较(0.352±0.018和0.220±0.012)、D组与B组比较(0.338±0.023和0.220±0.012),差异均有统计学意义(P<0.05)。

对组织石蜡切片进行免疫组化染色,分别检测每组标本中NF-κB p65 (p65)总蛋白的含量表达(图3A-D),为了方便观察p65蛋白在胞质及胞核内的表达情况,因此并未对切片进行胞核的苏木精染色。染色结果用Image J软件对切片染色进行平均吸光度值分析,结果显示,p65总蛋白主要在胞质内表达,LPS膀胱灌注2周及4周均能使大鼠膀胱上皮的p65总蛋白表达增加,与对照组间的差异有统计学意义(F=65.521,P<0.05 )。其中,A组与B组比较(0.208±0.025和0.198±0.025 ),差异无统计学意义(P=0.817>0.05);C组与B组比较(0.326±0.019和0.198±0.025),D组与B组比较(0.330±0.023和0.198±0.025 ),差异均有统计学意义(P<0.05)。
为了进一步观察p65蛋白的活化情况,对每组标本切片进行了磷酸化的NF-κB p65(以下简称NF-κB pp65)蛋白的免疫组化染色。结果显示(见图3E-H),pp65主要在胞核内表达,在胞质内也有少量表达,LPS膀胱灌注不仅能刺激大鼠膀胱上皮胞核内的pp65蛋白表达增加,还能使其在胞质内表达增加。染色结果用Image J软件分别对切片的胞核及胞质染色进行平均吸光度值分析,差异均有统计学意义(F=169.718 ,F=93.986,P<0.05)。对于胞核内pp65,其中,A组与B组比较(0.360±0.016和0.380±0.011),差异无统计学意义(P>0.05);C组与B组比较(0.475±0.019和0.380±0.011)、D组与B组进行组间比较(0.727±0.062和0.380± 0.011),差异均有统计学意义(P<0.05)。对于胞质内pp65,其中,A组与B组比较(0.182±0.010和0.193±0.006),差异无统计学意义(P>0.05);C组与B组比较(0.242±0.020和0.193±0.006)、D组与B组比较(0 . 334±0.029和0.193±0.006),差异均有统计学意义(P<0.05)。
对组织石蜡切片进行免疫组化染色,分别检测每组标本中NF-κB pp65的含量表达(图4A、图4B)。染色结果用Image J软件对切片染色结果进行AOD值分析,结果显示,PDTC能明显抑制pp65蛋白的表达,在胞核中,抑制剂组与对照组间的差异(0.449±0.032和0.721±0.035)有统计学意义(P<0.05 );在胞质中,抑制剂组与对照组间的差异(0.193±0.009和0.306±0.020)也有统计学意义(P<0.05)。
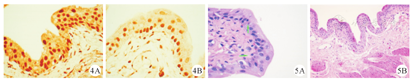
对HE染色病理切片进行镜下观察,结果显示,在E组大鼠膀胱中,仍可见散在的淋巴细胞在黏膜下层浸润。而在F组大鼠膀胱中,仍可见膀胱上皮的非典型增生,但未见明显的炎症细胞浸润。(图5A、图5B)
对每组大鼠实验终点时的6 h内的排尿次数进行了记录,结果提示:E组和F组大鼠6 h的排尿频率分别为(7.14±1.81)次、(4.25±1.56)次;对照组与抑制剂组比较,差异有统计学意义(t=3.097,P= 0.009<0.05)。
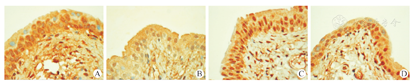
对组织切片进行Ⅰ型胶原纤维及Ⅲ型胶原纤维的免疫组化染色(图6A、图6B,图6C、图6D),AOD值结果提示:PDTC能一定程度上减少大鼠膀胱上皮中Col1蛋白的表达,抑制剂组与抑制剂对照组比较(0.242±0.017和0.333±0.027 ),差异有统计学意义(t=7.317,P<0.01);但其并不能明显减少大鼠膀胱上皮中Col3A1蛋白的表达,抑制剂组与抑制剂对照组比较(0.300±0.018和0.319± 0.016),差异无统计学意义(t=1.973,P>0.05)。
临床上存在下尿路功能障碍的患儿,容易出现泌尿系的反复感染,泌尿系感染导致的膀胱炎症作为膀胱纤维化的重要因素[11],有学者曾通过神经源性膀胱患儿的膀胱组织标本观察到膀胱壁中存在明显的炎性浸润、水肿及纤维化[12]。目前,较多学者通过动物膀胱炎症模型来研究膀胱纤维化的发生机制,大鼠的膀胱炎建模方法较多,如膀胱内灌注硫酸鱼精蛋白+LPS、盐酸、透明质酸酶或腹腔注射环磷酰胺等[13,14,15,16]。LPS是革兰阴性菌的主要致病因子,采用LPS诱导建立膀胱炎模型更能模拟临床上泌尿系感染所致膀胱炎的病理生理过程,该方法在既往研究中已被证实切实可行。Podmolikova等[4,5]通过膀胱内灌注LPS, LPS刺激使膀胱肥大细胞数量增加及促进炎症浸润,在LPS刺激14 d后膀胱炎症浸润更为明显,该方法成功诱导了大鼠膀胱炎模型的建立。膀胱纤维化是一种长期、慢性的病理过程,更长时间的建模处理将更有利于观察到膀胱胶原沉积的情况。在一些以纤维化和肥大细胞浸润为特征的间质性膀胱炎动物模型中,研究者们通过建模处理1个月左右即能观察到膀胱纤维化相关指标的变化[13,17]。因此本研究参考前人建模方法,使用LPS反复进行膀胱灌注来诱导大鼠膀胱炎模型,且为了更好地观察大鼠膀胱从炎症到发生纤维化的过程,本研究将建模处理时间延长至1个月。
本研究的实验结果显示:在LPS灌注2周左右,膀胱黏膜下层炎症浸润明显,主要为中性粒细胞,且伴随着膀胱上皮的非典型增生。此时,可能仍属于膀胱的急性炎症期。炎症依据时间长短分为急性炎症和慢性炎症。急性炎症持续时间短,一般不超过4周,浸润的炎症细胞主要为中性粒细胞。慢性炎症持续时间较长,数月到数年,以增生性病变为主,其浸润的炎症细胞主要为巨噬细胞及淋巴细胞[10]。本研究也再次证实了LPS膀胱灌注诱导大鼠膀胱炎建模方法的可行性。本实验将膀胱灌注的时间延长至4周,在LPS灌注4周后,我们观察到膀胱黏膜下层炎症浸润细胞已逐渐由中性粒细胞转变成了淋巴细胞,且伴随着黏膜上皮的糜烂、增生,表明膀胱炎症有转变为慢性炎症的趋势。
此外,由于膀胱炎症的刺激,B~F组大鼠均出现了不同程度的尿频症状。尿频作为最常见、也最容易观察到的下尿路刺激症状,在本研究中,观察到LPS灌注组大鼠的排尿频率均比对照组要高,尤其是在LPS灌注2周左右更为明显,虽然PBS灌注组较空白对照组也有一定程度的增加,其可能是反复导尿操作刺激所致。该结果也从临床症状上进一步证实了大鼠膀胱炎症模型的建立。
为了观察炎症刺激与纤维化之间的关系,本研究进一步检测了膀胱中两种胶原蛋白的表达情况,分别为:Ⅰ型胶原蛋白Col1和Ⅲ型胶原蛋白Col3的相关蛋白Col3A1。这两种类型的胶原蛋白是成纤维细胞分泌的主要胶原蛋白,对膀胱的生物力学功能起着至关重要的作用[18,19,20],且这两种类型的胶原蛋白的比例失衡也是导致膀胱顺应性下降的重要因素[21]。另一方面,细胞外基质的合成与降解之间的失衡是纤维化发生的关键,细胞外基质的过度沉积为膀胱纤维化的基本特征,而胶原蛋白作为细胞外基质的主要成分,其在膀胱中的过度分泌也一定程度上意味着纤维化进程的启动。在本研究观察到,这两种胶原蛋白不仅在间质中表达,在上皮细胞中也有表达;尤其是在LPS刺激后,这两种胶原蛋白在上皮细胞中的表达增加更为明显。由于免疫组化检测作为一种半定量半定性检测方法,因此本研究中并不能对这两种胶原蛋白的含量比例进行比较,这也是本研究的局限之处。
胶原蛋白作为细胞外基质的主要组成部分,主要由成纤维细胞合成分泌,其余在上皮细胞及内皮细胞中也有合成分泌。胶原纤维的基本组成成分为原胶原,主要在胞内合成,再分泌至间质中合成为粗纤维[18]。在本实验中,大鼠膀胱上皮反复直接暴露于LPS的刺激中,刺激细胞合成分泌胶原蛋白,这可能是上皮细胞比细胞间质的胶原蛋白表达更明显的原因。另一方面,随着人们对纤维化机制的认识,研究者们发现肌成纤维细胞并不完全来源于间质细胞,其可能是上皮细胞转化而来,人们把这个过程称为上皮-间质转化(epithelial-mesenchymal transition ,EMT ),这是一个指上皮细胞逐渐失去其功能和特性而获得间质细胞特性转化为间质细胞的过程[22,23,24]。EMT对于正常的伤口愈合是必要的,能促进上皮细胞及角质细胞失去细胞间粘附,向伤口迁移,并分泌合成细胞外基质促进创面愈合。上皮细胞发生EMT后,能分泌合成大量的细胞外基质,但假如上皮细胞向肌成纤维细胞的持续转化变得不可控就会导致纤维化疾病。Humphreys等[25]通过遗传谱系标记技术证实上皮细胞是肌成纤维细胞的重要来源,其他研究也报道了肺上皮细胞的反复损伤刺激导致的持续炎症反应及持续的EMT是导致肺纤维化的重要因素[26]。因此,在本研究中,这种对上皮细胞反复的刺激损伤,是否导致了上皮细胞发生EMT,从而使上皮细胞合成的胶原蛋白增多比间质细胞更明显?该假设需要后续进一步的研究来证实。
以上结果证实了LPS诱导的膀胱炎症可刺激膀胱上皮层胶原蛋白的合成增加,在此过程中,本研究检测了炎症通路——NF-κB信号通路相关蛋白的表达情况。NF-κB通路作为诱导炎症反应发生和发展的重要通路,该家族蛋白主要以二聚体的形式存在,而其信号通路的功能主要是依靠p50/p65这两者组成的二聚体来实现的。静息状态下,NF-κB二聚体与其抑制蛋白IκB形成一个三聚体复合物存在于细胞质中。IκB是NF-κB抑制蛋白,能屏蔽其NF-κB的核定位域,阻止NF-κB的激活,其成员包括IκBα、IκBβ、IκBε、IκBγ、Bcl3、IκBζ。NF-κB信号通路中另一个重要成员是IκB激酶(IKK),主要成员包括IKKα、IKKβ和IKKγ (NEMO )。当细胞受刺激后,多种信号分子如白细胞介素、肿瘤坏死因子-α(tumor necrosis factor α,TNF-α)、LPS等均能激活细胞内的NF-κB通路。信号分子与受体结合后,引起构象改变,激活IKK,IKK会发生磷酸化,与IκB结合使IκB发生磷酸化,磷酸化后的IκB会发生自身泛素化并裂解,释放出NF-κB二聚体,使其变成磷酸化的NF-κB,再转移入细胞核内与靶基因特定部位序列结合,启动基因的转录,产生更多的炎症介质并募集多种炎症细胞从而导致炎症的级联反应[27,28,29]。
关于NF-κB通路与LPS诱导的膀胱炎之间的关系,以前就有研究证实,使用LPS对小鼠进行膀胱灌注24 h后,膀胱可出现炎性特征,表现为黏膜溃疡、水肿、出血和炎症浸润,其中NF-κB通路是处于激活状态的[30]。在近来的研究中,Lai等[16]也在LPS处理过的人类膀胱平滑肌细胞中检测到活化的NF-κB信号分子。本研究通过更长时间的动物体内实验来探究该过程中NF-κB通路蛋白表达水平的变化,结果显示,在大鼠膀胱炎症刺激到胶原蛋白分泌增加的过程中,膀胱上皮中NF-κB p65总蛋白较对照组均有持续的高表达。为了进一步观察p65蛋白的活化情况,我们又对大鼠膀胱上皮进行了pp65的免疫组化检测,结果显示,不管在胞浆还是胞核中,pp65的含量均比对照组明显增高,尤其是在胞核中pp65的含量表达更高,说明炎症刺激胶原蛋白合成的过程中,NF-κB p65是在持续的激活。该结果绝非偶然,既往的研究结果证实,该通路在其他组织器官如肝脏、肺、肾脏等的纤维化的发生过程中也承担着重要作用[6,7,8],其可能机制是通过通路激活后产生的下游信号分子激活成纤维细胞,使其分泌的细胞外基质异常增多,造成细胞外基质在组织间的异常沉积及降解失衡,造成组织纤维化的发生。有研究结果报道[31],p65在肺的成纤维细胞中能通过转化生长因子-β(TGF-β)/Samd3通路进行表达上调,并协同调节TGF-β诱导肺的成纤维细胞的激活。因此,其可能作为TGF-β促纤维化信号通路上的信号分子的一员,也可能作为单独的促纤维化因子存在,目前仍需进一步的研究证实。
第一部分结果提示,LPS膀胱灌注可诱导大鼠膀胱炎症的发生,炎症刺激可诱导膀胱上皮的胶原分泌增加。在此过程中,NF-κB信号通路始终处于持续激活状态,NF-κB信号通路可能为炎症刺激胶原沉积过程中的重要信号通路,该通路的激活可能是纤维化过程的重要因素。这两者间的关系,仍需进一步的抑制剂实验来证实。
随着NF-κB通路在炎症、免疫等多种疾病的研究进展,多种NF-κB抑制剂已被研发出来,包括PDTC、菊内酯(parthenolide)、乳胞素(lactacystin)、姜黄素等[30,32]。PDTC目前在科研中应用较多,其作为一种抗氧化剂,能抑制IKK的活性从而抑制IκB的降解,进而抑制NF-κB的激活。Okabe等[9]证实,PDTC能通过减轻肾脏炎症反应进而减轻肾脏纤维化的发生。而在环磷酰胺诱导的小鼠膀胱炎/膀胱疼痛模型中,PDTC能明显减轻小鼠的膀胱疼痛症状[33]。因此本研究使用PDTC作为NF-kB抑制剂,在原有的大鼠膀胱炎模型中使用PDTC抑制NF-κB信号分子的活化,进一步观察NF-κB p65的活化与膀胱上皮细胞胶原分泌增加之间的相互关系,以及研究PDTC对膀胱上皮细胞胶原分泌的影响。
第二部分实验的结果显示,在使用PDTC后,大鼠膀胱上皮中胞核及胞质内的pp65的表达均收到了抑制,均比对照组要低。此时,抑制剂组大鼠膀胱的炎症细胞浸润程度以及尿频症状均比对照组大鼠有所好转,说明PDTC能一定程度上减轻大鼠膀胱的炎症程度以及大鼠的膀胱刺激症状。NF-κB信号通路作为诱导炎症反应的经典通路,在受到炎症细胞因子如IL-1、TNF-α及LPS等刺激下,可通过其"经典途径"激活NF-κB通路,调节促炎症因子的释放及炎症细胞的募集[3]。NF-κB通路也是淋巴细胞的存活、增殖所必需的,革兰阴性菌的内毒素LPS能通过刺激淋巴细胞上的Toll-like受体或与其表面上的IgM发生强交联而激活淋巴细胞[34]。当PDTC抑制NF-κB的激活后,可有效减轻淋巴细胞在膀胱黏膜下的浸润。PDTC应用于改善下尿路症状方面的相关报道较少。不过,Ozaki等[33]在环磷酰胺诱导的小鼠间质性膀胱炎/膀胱疼痛模型中,发现PDTC能明显减轻小鼠的膀胱疼痛症状。膀胱刺激症状与炎症程度相关,本实验中,PDTC可能是通过抑制NF-κB的激活,缓解膀胱炎症浸润,从而改善大鼠的尿频症状。
为了进一步观察PDTC抑制NF-κB通路激活后对膀胱纤维化的影响,本研究检测了膀胱中Col1和Col3两种胶原蛋白的表达情况。结果显示,抑制剂组大鼠膀胱上皮中Col1的含量比对照组明显降低,差异有统计学意义;Col3的含量抑制剂组虽比对照组有所降低,但差异并无统计学意义。结果说明,PDTC能在一定程度上抑制膀胱上皮中胶原蛋白的合成分泌,且以抑制Col1为主。目前,国内外关于PDTC用于治疗膀胱纤维化疾病方面的研究报道较为缺乏,但有研究[35]报道了PDTC在小鼠心肌梗死后心肌纤维化模型中,能有效抑制Col1的合成。
膀胱纤维化的一个主要特征就是细胞外基质的过度沉积导致膀胱顺应性的丧失,膀胱顺应性是指膀胱在充盈期维持其压力不变或仅轻度升高的能力,即膀胱对尿液增加的耐受力。胶原蛋白与弹性蛋白的比例对膀胱顺应性的作用至关重要,弹性蛋白比例升高,对膀胱的顺应性有利,而胶原蛋白的比例升高,则反之[36,37]。Col1和Col3这两种类型的胶原蛋白是成纤维细胞分泌的主要胶原蛋白,对膀胱的生物力学功能起着至关重要的作用[18,19,20]。Col1是最主要的间质胶原,可以合成厚纤维束,提供强大的抗张强度;Col1的异常增加会对平滑肌的顺应性造成负面影响,其在膀胱壁的过度沉积易致膀胱壁组织增厚、变硬、舒张能力下降。Col3作为第二主要的间质胶原,可以合成薄纤维束,为平滑肌提供弹性[36];其在膀胱壁的过度沉积易使膀胱的收缩能力降低。胶原蛋白在膀胱壁的异常沉积,易使膀胱壁组织增厚、变硬,最终发展成小容量、高压力的低顺应性膀胱[38]。另有研究表明[21],随着纤维化的进展,胶原蛋白的沉积在后期逐渐出现成份比例的改变,即非顺应性膀胱患儿的膀胱壁中Col3/Col1的比例较正常膀胱壁组织要高。在本实验中,使用PDTC后,胶原蛋白的总量有所下降,说明PDTC在早期能抑制膀胱上皮胶原蛋白的合成,能一定程度上延缓膀胱的纤维化进程。这种抑制作用主要以抑制Col1为主,对Col3的抑制作用并不明显,考虑可能:纤维化过程是一个复杂的病理过程,涉及多种信号通路,NF-κB炎症通路虽然被证实参与纤维化过程,但其并不作为纤维化的唯一信号通路。PDTC抑制该通路能一定程度上抑制细胞外基质的合成分泌,但其中仍可能有其他信号分子介导纤维化过程,抑制NF-κB通路并不能完全抑制所有类型的细胞外基质的合成分泌。研究表明[39],抑制TGFβ1受体、血小板源性生长因子受体以及血管内皮生长因子受体也能抑制Col3的合成。总的来说,膀胱纤维化都是细胞外基质在膀胱壁异常沉积的结果,各种类型的胶原蛋白对膀胱顺应性的作用都不可或缺。在本研究中,PDTC能在一定程度上抑制了膀胱上皮细胞中胶原蛋白的合成,这对于延缓膀胱纤维化的进程其实具有重要意义。但在研究抗纤维化机制时,仍需综合考虑其他的纤维化信号通路。
综上所述,PDTC作为一种NF-κB抑制剂,能有效抑制大鼠膀胱中NF-κB蛋白的活化,应用于LPS诱导的大鼠膀胱炎模型时,不仅能缓解大鼠膀胱的炎症程度以及尿频症状;还能在一定程度上抑制膀胱上皮细胞中胶原蛋白的合成分泌。NF-κB信号通路可能为炎症刺激膀胱发生纤维化过程中的重要通路,而抑制NF-κB通路的激活能一定程度上延缓膀胱纤维化的进程。
所有作者均声明不存在利益冲突

























